Biological Theory volume 14, pages141–150(2019) Cite this article
- Abstract
Anthropogenic changes in the biosphere, driven mainly by human cultural habits and technological advances, are altering the direction of evolution on Earth, with ongoing and permanent changes modifying uncountable interactions between organisms, the environment, and humankind itself. While numerous species may go extinct, others will be favored due to strong human influences. The Bio-Evolutionary Anthropocene hypothesizes that directly or indirectly human-driven organisms, including alien species, hybrids, and genetically modified organisms, will have major roles in the evolution of life on Earth, shifting the evolutionary pathways of all organisms through novel biological interactions in all habitats. We anticipate that, in future scenarios, novel organisms will be continuously created, and contemporary native organisms with no obvious economic use will decline—while anthropogenic-favored and novel organisms will spread. The Bio-Evolutionary Anthropocene hypothesis therefore predicts that humankind and novel organisms will interact within a strong evolutionary bias that will lead to unexpected, and probably irreversible, outcomes for the evolution of life on our planet.
Human Hyper-Dominance Leading to Changes
Human beings have drastically impacted the Earth’s surface and promoted striking ecosystem and biodiversity alterations (Ehrlich and Holdren 1971; Vitousek 1997; Chapin et al. 2000; Butchart et al. 2010; Steffen et al. 2011). Habitat destruction and pollution, species extinctions, biotic homogenization (McKinney and Lockwood 1999), and gene exchange between species (Bawa and Anilakumar 2013) are some of the many ways the biosphere is changing. On the other hand, despite the apparent biological impoverishment of Earth’s ecosystems, humans could actually be directly increasing biodiversity (McClure 2013; Thomas 2013; Fuentes 2018), as anthropogenic ecosystems, such as cities, may drive evolution and create new organisms (Johnson and Munshi-South 2017)—thus establishing new evolutionary pathways created by human hyper-dominance as a “hyper-keystone” species (Worm and Paine 2016).
These contradictory views of anthropogenic influences divide scientific opinion about whether human-induced changes are positive, as many organisms are favored by artificial selection, or if Earth is nearing its sixth mass extinction (Dalby 2016). The Anthropocene is surely a time of mass disruptive processes on a planet that has already been fundamentally altered by humans (Hamilton 2016), while population pressures on Earth’s ecosystems are exponentially increasing (Deb et al. 2018). It is therefore irrefutable that species distributions, species richness, and novel organisms will diverge enormously from contemporary biodiversity in the Anthropocene. Human cultural values and other social structures lead to behavioral patterns (of both individuals and social groups) that will result in drastic environmental changes (Ellis and Trachtenberg 2014). Some anthropogenic effects are now easily visible, such as habitat extinction (Ghosh et al. 2013) and the production of ~ 30 trillion tons (Tt) of technosphere materials and artifacts (Zalasiewicz et al. 2017). There are, however, numerous less-noticeable environmental impacts, including the high production of pesticides (Chagnon et al. 2015), fertilizers (Vitousek et al. 2009), acidic effluents (Akcil and Koldas 2006), radioactive wastes (Geraskin et al. 2003), antimicrobial compounds (Gillings and Stokes 2012), the spread of genetically modified organisms (GMOs) (Bawa and Anilakumar 2013), and the spread of alien species (Lodge 1993; Simberloff et al. 2013)—especially within or near cities (McKinney 2006; Toussaint et al. 2016).
Those modifications greatly disturb natural ecosystems, although certain organisms will be favored based on their capacity to adapt to the new anthropogenic conditions, resulting in the emergence of novel and better-adapted organisms that will become established and persist in modified areas or habitats. Some of the areas altered by humans are considered new ecosystems—known as anthropogenic biomes (e.g., anthromes; Ellis 2011).
The Anthropocene is modulated by human culture and technology, and extinctions and habitat changes are occurring at uncontrolled and accelerated rates with unexpected consequences (Barnosky et al. 2012; Steffen et al. 2015). This trend can be confused with the “tipping point” hypothesis, which argues that evolutionary patterns are permanently changed by anthropogenic pressures and biological thresholds are definitely crossed (Hodgson et al. 2015; van Nes et al. 2016). The Bio-Evolutionary Anthropocene hypothesis, as explained here, however, recognizes novel organisms and humankind as the new driving forces of biodiversity. The future of biodiversity is hardly predictable with any precision, of course, as many global characteristics such as functional diversity, novel organisms, and atmospheric pollution cannot yet be fully factored into a future vision (Steffen et al. 2007, 2015).
We anticipate that novel organisms, such as alien and hybrid species and GMOs will play key roles in biological interactions—leading to what we call the Bio-Evolutionary Anthropocene. Those organisms will have divergent evolutionary capacities or create different pressures on both natural and anthropized ecosystems and alter the distribution, richness, and ecological patterns of local and global biodiversity—and lead to novel and unexpected evolutionary pathways. We will discuss here the roles of those novel organisms on biodiversity and evolution and the resulting consequences for the biosphere from the perspective of the Bio-Evolutionary Anthropocene hypothesis.
The Bio-Evolutionary Anthropocene Hypothesis
The Bio-Evolutionary Anthropocene hypothesis is largely based on the emergence and establishment of novel organisms and their new biological interactions in natural or modified habitats. The creation, spread, and transformation of new life forms are mainly induced and favored by human activities, and therefore represent the most important human imprints on evolution.
Evolution is a modifying, transforming, and changeable force that reflects the interactions of species (Thompson 1999). Evolutionary pathways are not static, but constantly changing due to direct or indirect human interference (Otto 2018; Pelletier and Coltman 2018) and impacts on biological interactions. Those changes, either driven by, or the random results of human actions, are immediately imprinted on all living organisms through habitat modifications, novel or lost functions, and new interactions (Morse et al. 2014; Pigeon et al. 2016; Rudman et al. 2017)—eventually leading to a “point of no return” when those changes become permanent (sensu Corlett 2015).
According to the Modern Synthesis, while natural selection, genetic drift, gene flow, and mutation have been considered the principal evolutionary forces (Charlesworth et al. 2017), other processes, such as developmental bias, plasticity, inclusive inheritance, and niche construction can also contribute to species evolution (Laland et al. 2015). That perspective incorporates both natural processes and those modulated by humans, thus consolidating the role of humans in biological evolution.
The evolutionary shifts that are occurring now in the biosphere are harsher and faster than previously expected (Otto 2018)—especially if we consider the emergence and spread of novel organisms (mainly alien species, hybrids, and GMOs) that will continue to be produced and spread by humans (either purposely or accidentally). The consequences of the interactions of those novel organisms with contemporary ones, added to constantly changing environments in the Anthropocene, are not fully predictable. In many ways and intensities, humans are modulating and pushing new evolutionary outcomes towards the Bio-Evolutionary Anthropocene in uncountable ways and directions.
The Bio-Evolutionary Anthropocene hypothesis therefore incorporates multi-scaled and fractal changes in additive biodiversity patterns, inevitably shifting local and global evolutionary pathways. Future biodiversity scenarios (modified ecosystems derived from the Biological Anthropocene hypothesis) will likely demand that organisms adapt more and more rapidly (than they would normally) in response to rapid ecosystem changes. Some examples of current modified ecosystems created and modulated by humans are: semi-natural habitats (e.g., Kalusová et al. 2017), agricultural fields (Vanwalleghem et al. 2017), anthropogenic biomes (Ellis 2011), and urban (Alberti 2015) and novel ecosystems (Morse et al. 2014).
Because of current and future human evolutionary pressures, the Bio-Evolutionary Anthropocene hypothesis predicts that organisms capable of fast adaptation to new or modified habitats, such as alien species, hybrids, GMOs, and economically and anthropogenically favored organisms (e.g., crops and livestock) will prevail. Indeed, those organisms will not only persist, but be favored in increasingly modified environments because of their resilience and high capacity for adaptation.
The strong influence of humans over new or transformed biological entities (now, and in the future) in the Bio-Evolutionary Anthropocene hypothesis is mainly driven by human-related processes including: induced or produced hybridizations (e.g., botanical gardens and ornamental flowers), artificial selection (e.g., crop domestication and improvement), positive selection (e.g., plagues and parasites), environmental transformations, alien species establishment, and the spread and exchange of modified genes through biotechnology. In our hypothesis, these and other related processes will modulate the Anthropocene in feedback loops—with uncountable new interactions between organisms, humans themselves, and human cultures and technologies.
In the Bio-Evolutionary Anthropocene hypothesis, we understand that humans are the main driving force—not only creating but also linking human-driven processes with modified habitats, and novel and contemporary organisms. Therefore, humans not only induce changes, but will be one of the main species affected by anthropogenic changes—transforming and simultaneously being transformed. Those interactive loops are definitely reshaping and shifting evolutionary pathways and Earth’s biodiversity in unprecedented ways (Fig. 1).
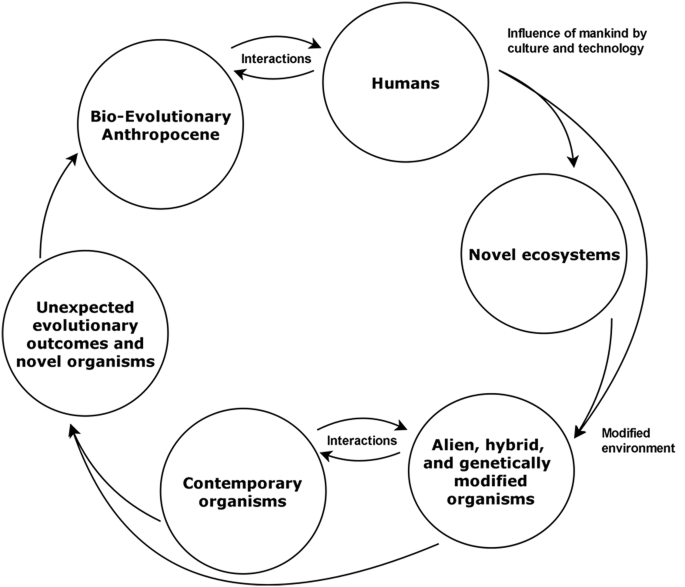
The Anthropocene: Facts and Mechanisms
Human hyper-dominance is the main driving force of the Anthropocene. Humans, even as a single species, have completely changed the environment and evolutionary outcomes through hyper-dominance, technological development, and diverse cultural habits (Worm and Paine 2016). Human–environment interactions are not just local anymore, but have spread to ever widening spatial scales (Steffen et al. 2015; Sullivan et al. 2017), largely due to human technological advances, and social and cultural values (Alberti 2015).
This new concept of the human–environment relationship, incorporating mankind’s advances throughout the world, can be easily perceived, but general awareness of that situation is still incipient. It is currently difficult to identify environmental modifications and species interactions that are not driven, intentionally or not, by human culture and technology (Lewis and Maslin 2015). Humans occupy almost the entire planet, without any special habitat distinctions, so that essentially all areas have in some way been impacted by humans (Clark 1996).
Human interventions in the environment have lead to many uncertainties and risks, whose consequences are sometimes negative and sometimes positive. The Bio-Evolutionary Anthropocene hypothesis incorporates the concept that human-influenced organisms can permanently modify biological evolution. While still unpredictable, there are strong signs that the new patterns predicted by the new Bio-Evolutionary Anthropocene hypothesis are already becoming established.
Even conservation efforts as we know them today, for example, could result in unexpected outcomes that alter natural evolutionary processes. Intentional conservation actions favoring a few selected organisms (such as flagship species) change population patterns within an ecosystem (Liu et al. 2007), consequently even cosmopolitan species are similarly experiencing habitat losses and modifications (Simmonds et al. 2019). Future outcomes predicted by modeling studies may never come about because of the constant and unpredictable interactions between living organisms and their environments, especially in the Anthropocene.
The patterns and processes predicted by the Bio-Evolutionary Anthropocene hypothesis can be viewed as having started thousands of years ago as, from the times of our ancestors (from Neanderthals to modern Homo sapiens), all anthropogenic actions, decisions, necessities, and even ideas have altered the outcome of evolution (see the “Early Anthropogenic hypothesis”; Rudman et al. 2017). Some examples of current anthropogenic impacts leading to biodiversity instability and unexpected outcomes are: species exploitation (hunting or collecting) (Otto 2018), trophic cascades and predator–prey interactions (Allan et al. 2013; Dorresteijn et al. 2015), pollinator population declines (Potts et al. 2010), and the spread of parasite vectors (Civitello et al. 2015).
Population declines, species extinctions, and habitat losses have already caused irreversible changes in the dynamics of most ecosystems and entire assemblages of both common and threatened species (Simmonds et al. 2019). Those human-driven changes will eventually affect species all over the globe. Our current scientific knowledge is much too underdeveloped to fully understand or predict the spectrum of changes in species interactions that will occur due to our social and cultural habits and long-term technological progress.
In addition to changes and environmental shifts caused by human actions, the Bio-Evolutionary Anthropocene hypothesis considers novel organisms created directly or indirectly by human-driven processes such as: artificial selection (Allendorf and Hard 2009; Driscoll et al. 2009; Otto 2018), hybridization (Mallet 2005), ploidy changes in animals (Otto 2007) and plants (De Storme and Mason 2014), as well as transgenic organisms (Bawa and Anilakumar 2013). Additionally, many changes in species compositions (biotic) (Stephens et al. 2009) and environmental conditions (abiotic) have been influenced by human actions (Bull and Maron 2016; Hendry et al. 2017; Nadeau et al. 2017).
In the Bio-Evolutionary Anthropocene hypothesis, although biotic and abiotic alterations of habitat are usually pitfalls for contemporary organisms, they could favor the establishment of novel organisms in modified habitats. The main novelties we considered here are alien species, hybrids, and genetically modified organisms that are currently integrating new webs of interactions in semi-natural or highly modified habitats.
Alien organisms first began to spread due to different human processes: the domestication of plants and animals, farming and animal husbandry, urban planning and landscaping, and many others. Those organisms were, and still are, being modified from ancestral species or similar varieties, transported to different and modified environments, and established or cultivated by humans based on biological necessities, cultural and social traditions, and technological advances.
Nowadays, alien organisms interfere with and redefine biotic and abiotic conditions in many anthropogenic-influenced habitats, especially due to their invasiveness, resilience, and high capacity for adaptation and rapid evolutionary alterations (see Cox 2004). Many efforts focusing on the eradication of alien species are inefficient because of their high fitness and resilience (Pimentel et al. 2001). Alien organisms will therefore definitely be present—even abundant—in many human-modified ecosystems according to the Bio-Evolutionary Anthropocene hypothesis.
Human-induced hybrids are another form of novel organisms condemned by many conservationists as a threat to parental species integrity (Rhymer and Simberloff 1996), but we predict that some of them will thrive in the Anthropocene. The propagation of hybrids in modified ecosystems could cause the decline of parental species, and those processes are therefore considered unnatural and in need of ecosystem management (Muhlfeld et al. 2014). On the other hand, there are many examples of human-induced hybrids that are positive and successful (Grant and Grant 1992; Huxel 1999), and they may change future evolutionary outcomes.
The third novelty addressed here concerns genetically modified organisms. International debates concerning GMOs take two opposite perspectives, although both have anthropocentric points of view in common. While supporters believe that humanity needs these organisms for food security, critics maintain that there are many uncertainties about environmental and health risks (Wolfenbarger and Phifer 2000; Ellstrand et al. 2013). Even now, genetic modifications are present in almost all food crops (Zhang et al. 2016), with the annual cultivation of billions of transgenic organisms in permeable anthropogenic ecosystems that frequently allow interactions with contemporary organisms. Besides traditional transgenic organisms, other GMOs are emerging through technological and scientific advances such as cisgenic plants and epicrops. Those organisms will also likely influence evolutionary pathways in the future and confirm the strong influence of humans’ biotechnology and the creation and establishment of novel organisms in all habitats.
In addition to the novel organisms cited here, the Bio-Evolutionary Anthropocene hypothesis predicts that others could unexpectedly emerge due to interactions between organisms, human influences, environment modifications, new technological advances, and novel human cultural and social habits. The processes and mechanisms incorporated into the Bio-Evolutionary Anthropocene hypothesis could favor either contemporary or novel organisms, and all of the possible consequences of those already shifting evolutionary pathways cannot be anticipated. Interactive feedback loops are additive, according to the Bio-Evolutionary Anthropocene hypothesis, and mostly unexpected, leading to an uncertain future biota.
Alien Organisms
It is common sense that alien organisms (non-native to a given habitat or ecosystem) represent real threats to local biodiversity once they become established and widespread, usually due to (intentional or unintentional) human actions (Walther et al. 2009). In highly anthropized and changing ecosystems, however, alien species can become “survivors” if they are directly favored by human activities such as artificial selection and domestication (e.g., Milla et al. 2015). In that sense, even crop plants and livestock are alien organisms that are intentionally farmed and raised in non-native areas. Many studies have shown that alien species not only colonize new habitats but also modify them (e.g., Elton 1958; Pimentel et al. 2001; Cox 2004) and can reduce local biodiversity (Lodge 1993; Simberloff et al. 2013). Alien organisms generally spread easily as a result of human activities (Richardson et al. 2000; Pimentel et al. 2001), and as humans are widespread on Earth, alien species have reached essentially every corner of the planet, affecting and modifying the environment as intensely as past mass extinctions (Barnosky et al. 2011)—resulting in severe biotic homogenization that may lead to pools of species similar to those within anthropized habitats (Lodge 1993; McKinney and Lockwood 1999).
The fact that alien organisms survive and spread in invaded habitats, provoking the decimation of native organisms, shows that those novel organisms are more suitable to, and apparently favored by, their new habitat—usually due to their high adaptability and reproductive capacities. Many newly invaded habitats show large alien populations, mostly in highly anthropized sites, which are difficult to manage. Although pristine areas seems to be less vulnerable to biotic invasions (e.g., Foxcroft et al. 2011), alien organisms thrive even in legally protected areas. In the Bio-Evolutionary Anthropocene hypothesis, alien organisms represent not only a disruption of natural processes, but also a new way to resist and even benefit from human-driven changes. Those organisms can take on key roles in biotic and abiotic interactions, especially in already modified habitats.
Hybrid Organisms
Hybridization is an important evolutionary force for species diversification (Mallet 2005), but it can be driven by humans, whether intentionally or not, in the Anthropocene. The creation of hybrid organisms can be positive when it results in species diversification or domestication, or negative when fostering or diversifying disease agents and vectors or pests (Arnold 2004; Ellstrand et al. 2013). The process of hybrid speciation has been found to be much more common in nature than previously thought, in terms of both plants and animals, so that hybridization may also catalyze major evolutionary innovations (Mallet 2007). Hybridization among species has been boosted by human technology and altered habits that open new possibilities for speciation. Horizontal gene transfer, for example, a mechanism by which unrelated species share genetic material, is a potential asexual mechanism for plant evolution and speciation through the commonly used technique of grafting (Fuentes et al. 2014). Since successful horizontal transfer of genetic material between unrelated species usually favors the development of novel traits, it may increase the adaptive capacities of those hybrid organisms and thus shape evolution (Soucy et al. 2015).
Hybrid speciation also occurs through genetic admixtures (sexual mechanisms). Those events are common in plants but thought to be rare in vertebrates. Accumulating evidence, however, indicates the contrary, with numerous examples of successfully adapted hybrid animal species that highlight the importance of hybridization as a source of genetic variation for speciation processes and as a source of evolutionary novelties (Barrera-Guzmán et al. 2017). Most successfully established hybrids have higher fitnesses than their parental organisms (Crispo et al. 2011). Even if their fitness is not high, models have shown that hybrids can naturally and rapidly evolve into new species through reproductive isolation driven by genetic incompatibilities (Schumer et al. 2015). Additionally, hybrids may evolve differently in anthropogenic-related or natural hybrid zones, as seen with monkey hybrids (Callithrix) in Brazil. Those hybrids are genetically differentiated in anthropogenically impacted areas (such as urban ecosystems), and scientists cannot accurately anticipate the futures of either the hybrids or their parental species (Malukiewicz et al. 2015).
Briefly, hybridization is a strong evolutionary force adding diversity and adaptive capacities to both old and new species, and it has a much greater role in evolution than previously recognized—as hybrids usually outperform parental species in altered habitats (Chunco 2014). Hybrids induced by anthropogenic actions seem to be strongly altering the current balance of biodiversity and successfully adapting to both novel and altered ecosystems. As such, hybrid organisms will likely become common in anthropized areas where their parent species have gone extinct. Silent hybridization mechanisms, such as horizontal gene transfer and introgression, may also generate new traits in changing environments, quickly transforming current species richness and distributions.
Genetically Modified Organisms (GMOs)
Human-induced gene exchanges between organisms may be one of the strongest life-changing mechanisms on Earth. The broad definition of GMO includes transgenic organisms (where part of the genetic material of one species is transferred to another) (Zhang et al. 2016); cisgenic organisms (that have introduced genes originally from the same or a sexually compatible species) (Kost et al. 2015); and, more recently, epicrops (which have undergone epigenetic alterations involving agronomically important traits) (Song et al. 2017). We will focus here on the most studied type of GMO, transgenic organisms which are possibly the most controversial organisms yet directly created to satisfy human needs.
Transgenic organisms favor humans directly by providing, for example, vaccines and drugs (Ma et al. 2005), but they may herald unexpected or undesirable outcomes—such as the spread of new pests (Cheke 2018) and increased mortality in non-target species (Losey et al. 1999). GMO crops are largely produced in developed countries (and some developing countries, such as Argentina, Brazil, China, and India), due to their high yields and low labor requirements as compared to organic farming—in spite of their high potential for environmental and health risks (Azadi and Ho 2010).
Although there are many uncertainties about the risks of transgene spread that could be caused by hybridization and introgression events in areas bordering agrosystems (Ellstrand et al. 2013), transgenic hybrids could have higher fitnesses than their parental organisms (Dong et al. 2017). Although many believe that biotechnological advances are essential to rapidly creating new strains of GMO crops to achieve sustainable global food security (Zhang et al. 2016), GMO crops may also alter natural processes and functions in the ecosystems around them (Catarino et al. 2015).
Similarly, the success of gene-edited animals (to correct genetic defects or increase disease resistance) (Van Eenennaam 2017) and scientific advances in terms of genetically modified livestock and fish (for food or feed production) (Forabosco et al. 2013), will likely result in the creation and emergence of more novel organisms in the near future. As such, even in light of the uncertain consequences of the global spread of crop and animal GMOs, there is enormous pressure to create and produce novel, better-adapted genetically modified crops and livestock strains through modern biotechnological techniques to supply human needs.
The constant development and improvements of technologies to genetically modify organisms will generate even more uncertainties as GMOs spread. There will certainly be more novel organisms in the future linked to anthropogenic changes—transforming evolution along unexpected and unforeseen paths.
Theoretical Scenarios
Changes in the landscape, organisms, humans, and habits will result in new and unexpected scenarios due to novel organisms and their interactions. So, in spite of the obvious stochasticity and unpredictability of biodiversity in the Anthropocene, evolution will certainly be reshaped by human actions. In order to represent the Bio-Evolutionary Anthropocene hypothesis schematically, we present here general hypothetical scenarios concerning novel organisms for the near present as well as three possible future scenarios.
In this approach, we grouped living organisms as native, alien, or anthropogenically favored organisms. Native organisms are the contemporary wild organism in natural ecosystems; aliens represent all non-native organisms in natural or semi-natural ecosystems; anthropogenic-favored organisms are urban, crop plants, livestock, or other organisms favored by human actions. Two variables based on human-driven changes on Earth were used for scenario constructions to determine the expansion or retraction of organism distributions based on the likelihood of their success and establishment, versus population reduction and extinction, according to the Bio-Evolutionary Anthropocene hypothesis. The variables considered were: environmental degradation and climate change (indirect human-driven changes), and human expansion and the use of natural resources (direct human-driven changes).
Considering those hypothetical scenarios, the more intense indirect human-driven changes on global climate and environments are (x-axis, Fig. 2), the more alien organisms will spread; while anthropogenic-favored organisms will spread more with increasing direct influence of humans on environments and organisms (y-axis, Fig. 2). Based on the Bio-Evolutionary Anthropocene hypothesis, alien organisms (which have high adaptive capacities), will likely advance into modified habitats, while contemporary native organism populations will become reduced due to competition or their poor adaptive capacities.
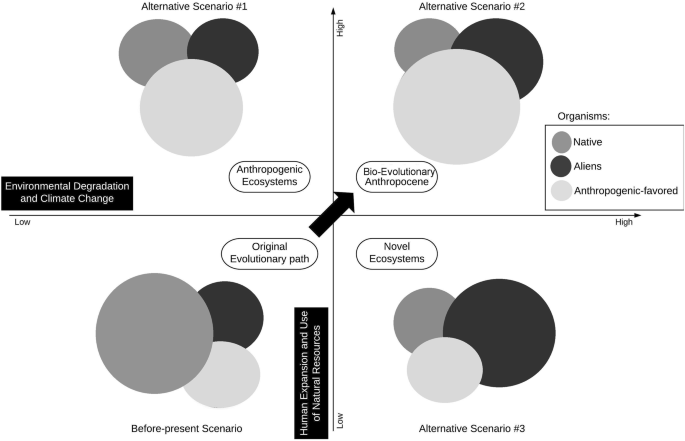
Anthropogenic-favored organisms, i.e., organisms not only adapted to urban or anthropized habitats but also created and/or positively favored by human actions—such as plagues, disease-vectors and agents, plants used for landscaping, economically favored species, hybrids, transgenics, and other associated organisms—will mostly expand when there is high direct human interference on the environment, whereas native organisms will suffer due to habitat and population reductions and possibly go extinct.
In all three predicted scenarios, native organisms will suffer reductions due to competition, habitat degradation or modification, and other reasons. This outcome for native organisms, although apparently very drastic, is very plausible in the near future. The Bio-Evolutionary Anthropocene hypothesis anticipates that fate for native organisms due to the view that alien, hybrid, genetically modified organisms, and all anthropogenically favored organisms will reshape evolution on this planet, shifting pathways by direct and indirect human actions without the possibility of mitigation.
The proposed scenarios are simplifications of complex cause-and-effect relationships and show possible outcomes based on the increase or mitigation of human-driven modifications, according to the Bio-Evolutionary Anthropocene hypothesis. Interestingly, the three hypothetical scenarios can be related to alternative ecological scenarios: #1 (top left) relates to anthropogenic ecosystem theory; #2 (top right) relates to the Bio-Evolutionary Anthropocene hypothesis; and, #3 (bottom right) depicts a novel ecosystem concept (Fig. 2).
The Bio-Evolutionary Anthropocene hypothesis is represented as alternative scenario #2, where both variables reach high levels. Since human-driven changes on Earth are possibly irreparable, unexpected outcomes of the Anthropocene are imminent. In that scenario, environmental degradation and climate modifications are very high and beyond repair, and direct human-driven modifications, such as the use of natural resources through human hyper-dominance and expansion, are immeasurable and astonishingly high, so that evolutionary pathways will be shifted permanently, with emerging novel organisms and novel interactions among them, humans, and the modified environment.
Briefly, future ecosystems and environments will have little space for contemporary native organisms with no economic use, and they may become extinct due to human-driven changes. Alien organisms and anthropogenic-favored organisms, on the other hand, will emerge and spread in disorderly ways, creating novel interactions and future novel organisms along unexpected evolutionary pathways. Although the Bio-Evolutionary Anthropocene hypothesis might seem negative and unwanted, it is quite plausible and has unavoidable outcomes. Global awareness must increase, with the realization that humans now represent the most important evolutionary force, and all future predictions must incorporate the processes and mechanisms inherent in every human action.
Conclusions
Humans are altering the path of evolution and the surface of the earth in unprecedented ways. Technological advances and the hyper-dominance of humans have created new habitats and novel evolutionary pressures on all organisms, and will lead to huge biodiversity losses. Some anthropogenic changes, however, including the emergence of novel organisms (constantly introduced and established in natural and semi-natural ecosystems), can be interpreted as a type of new adaptable biodiversity that will reshape evolution on Earth. According to the Bio-Evolutionary Anthropocene hypothesis, those novel organisms are alien and hybrid species, GMOs, and other organisms either created or induced by humans based on our habits, cultures, and technologies. Interactions between all organisms constantly change and adapt by additive and fractal patterns, leading to unexpected ecological outcomes. This new hypothesis therefore considers humans and novel organisms as key components of evolution and, even in controlled environments, unforeseen interactions will likely occur resulting in unexpected future scenarios and outcomes.
References
- Akcil A, Koldas S (2006) Acid mine drainage (AMD): causes, treatment and case studies. J Clean Prod 14:1139–1145. https://doi.org/10.1016/j.jclepro.2004.09.006Article Google Scholar
- Alberti M (2015) Eco-evolutionary dynamics in an urbanizing planet. Trends Ecol Evol 30:114–126. https://doi.org/10.1016/j.tree.2014.11.007Article Google Scholar
- Allan BJM, Domenici P, McCormick MI, Watson S-A, Munday PL (2013) Elevated CO2 affects predator-prey interactions through altered performance. PLoS ONE 8:e58520. https://doi.org/10.1371/journal.pone.0058520Article Google Scholar
- Allendorf FW, Hard JJ (2009) Human-induced evolution caused by unnatural selection through harvest of wild animals. Proc Natl Acad Sci USA 106:9987–9994. https://doi.org/10.1073/pnas.0901069106Article Google Scholar
- Arnold ML (2004) Natural hybridization and the evolution of domesticated, pest and disease organisms. Mol Ecol 13:997–1007. https://doi.org/10.1111/j.1365-294X.2004.02145.xArticle Google Scholar
- Azadi H, Ho P (2010) Genetically modified and organic crops in developing countries: a review of options for food security. Biotechnol Adv 28:160–168Article Google Scholar
- Barnosky AD, Matzke N, Tomiya S, Wogan GOU, Swartz B et al (2011) Has the earth’s sixth mass extinction already arrived? Nature 471:51–57. https://doi.org/10.1038/nature09678Article Google Scholar
- Barnosky AD, Hadly EA, Bascompte J, Berlow EL, Brown JH et al (2012) Approaching a state shift in Earth’s biosphere. Nature 486:52–58. https://doi.org/10.1038/nature11018Article Google Scholar
- Barrera-Guzmán AO, Aleixo A, Shawkey MD, Weir JT (2017) Hybrid speciation leads to novel male secondary sexual ornamentation of an Amazonian bird. Proc Natl Acad Sci USA 115:E218–E225. https://doi.org/10.1073/pnas.1717319115Article Google Scholar
- Bawa AS, Anilakumar KR (2013) Genetically modified foods: safety, risks and public concerns—a review. J Food Sci Technol 50:1035–1046. https://doi.org/10.1007/s13197-012-0899-1Article Google Scholar
- Bull JW, Maron M (2016) How humans drive speciation as well as extinction. Proc R Soc B 283:20160600. https://doi.org/10.1098/rspb.2016.0600Article Google Scholar
- Butchart SHM, Walpole M, Collen B, van Strien A, Scharlemann JP et al (2010) Global biodiversity: indicators of recent declines. Science 328:1164–1168. https://doi.org/10.1126/science.1187512Article Google Scholar
- Catarino R, Ceddia G, Areal FJ, Park J (2015) The impact of secondary pests on Bacillus thuringiensis (Bt) crops. Plant Biotechnol J 13:601–612. https://doi.org/10.1111/pbi.12363Article Google Scholar
- Chagnon M, Kreutzweiser D, Mitchell EAD, Morrissey CA, Noome DA, Van der Sluijs JP (2015) Risks of large-scale use of systemic insecticides to ecosystem functioning and services. Environ Sci Pollut Res 22:119–134. https://doi.org/10.1007/s11356-014-3277-xArticle Google Scholar
- Chapin FS III, Zavaleta ES, Eviner VT, Naylor RL, Vitousek PM et al (2000) Consequences of changing biodiversity. Nature 405:234–242. https://doi.org/10.1038/35012241Article Google Scholar
- Charlesworth D, Barton NH, Charlesworth B (2017) The sources of adaptive variation. Proc R Soc B 284:20162864. https://doi.org/10.1098/rspb.2016.2864Article Google Scholar
- Cheke RA (2018) New pests for old as GMOs bring on substitute pests. Proc Natl Acad Sci USA 115:8239–8240. https://doi.org/10.1073/pnas.1811261115Article Google Scholar
- Chunco AJ (2014) Hybridization in a warmer world. Ecol Evol 4:2019–2031Article Google Scholar
- Civitello DJ, Cohen J, Fatima H, Halstead NT, Liriano J et al (2015) Biodiversity inhibits parasites: broad evidence for the dilution effect. Proc Natl Acad Sci USA 112:8667–8671. https://doi.org/10.1073/pnas.1506279112Article Google Scholar
- Clark DB (1996) Abolishing virginity. J Trop Ecol 12:735–739. https://doi.org/10.1017/S0266467400009937Article Google Scholar
- Corlett RT (2015) The Anthropocene concept in ecology and conservation. Trends Ecol Evol 30:36–41. https://doi.org/10.1016/j.tree.2014.10.007Article Google Scholar
- Cox GW (2004) Alien species and evolution: the evolutionary ecology of exotic plants, animals, microbes, and interacting native species. Nature 432:276–277Article Google Scholar
- Crispo E, Moore J-S, Lee-Yaw JA, Gray SM, Haller BC (2011) Broken barriers: human-induced changes to gene flow and introgression in animals. BioEssays 33:508–518. https://doi.org/10.1002/bies.201000154Article Google Scholar
- Dalby S (2016) Framing the Anthropocene: the good, the bad and the ugly. Anthr Rev 3:33–51. https://doi.org/10.1177/2053019615618681Article Google Scholar
- De Storme N, Mason A (2014) Plant speciation through chromosome instability and ploidy change: cellular mechanisms, molecular factors and evolutionary relevance. Curr Plant Biol 1:10–33. https://doi.org/10.1016/j.cpb.2014.09.002Article Google Scholar
- Deb S, Debnath MK, Chakraborty S, Weindorf DC, Kumar D et al (2018) Anthropogenic impacts on forest land use and land cover change: modelling future possibilities in the Himalayan Terai. Anthropocene 21:32–41. https://doi.org/10.1016/j.ancene.2018.01.001Article Google Scholar
- Dong SS, Xiao MQ, Ouyang DX, Rong J, Lu B-R et al (2017) Persistence of transgenes in wild rice populations depends on the interaction between genetic background of recipients and environmental conditions. Ann Appl Biol 171:202–213. https://doi.org/10.1111/aab.12365Article Google Scholar
- Dorresteijn I, Schultner J, Nimmo DG, Fischer J, Hanspach J et al (2015) Incorporating anthropogenic effects into trophic ecology: predator–prey interactions in a human-dominated landscape. Proc R Soc B 282:20151602. https://doi.org/10.1098/rspb.2015.1602Article Google Scholar
- Driscoll CA, Macdonald DW, O’Brien SJ (2009) From wild animals to domestic pets, an evolutionary view of domestication. Proc Natl Acad Sci USA 106:9971–9978. https://doi.org/10.1073/pnas.0901586106Article Google Scholar
- Ehrlich PR, Holdren JP (1971) Impact of population growth. Science 171:1212–1217. https://doi.org/10.1126/science.171.3977.1212Article Google Scholar
- Ellis EC (2011) Anthropogenic transformation of the terrestrial biosphere. Philos Trans R Soc A 369:1010–1035. https://doi.org/10.1098/rsta.2010.0331Article Google Scholar
- Ellis MA, Trachtenberg Z (2014) Which Anthropocene is it to be? Beyond geology to a moral and public discourse. Earth’s Future 2:122–125. https://doi.org/10.1002/2013EF000191Article Google Scholar
- Ellstrand NC, Meirmans P, Rong J, Bartsch D, Ghosh A et al (2013) Introgression of crop alleles into wild or weedy populations. Annu Rev Ecol Evol Syst 44:325–345. https://doi.org/10.1146/annurev-ecolsys-110512-135840Article Google Scholar
- Elton CS (1958) The ecology of invasions by animals and plants. Springer, BostonGoogle Scholar
- Forabosco F, Löhmus M, Rydhmer L, Sundström LF (2013) Genetically modified farm animals and fish in agriculture: a review. Livest Sci 153:1–9. https://doi.org/10.1016/j.livsci.2013.01.002Article Google Scholar
- Foxcroft LC, Jarošík V, Pyšek P, Richardson DM, Rouget M (2011) Protected-area boundaries as filters of plant invasions. Conserv Biol 25:400–405. https://doi.org/10.1111/j.1523-1739.2010.01617.xArticle Google Scholar
- Fuentes M (2018) Biological novelty in the anthropocene. J Theor Biol 437:137–140. https://doi.org/10.1016/j.jtbi.2017.10.027Article Google Scholar
- Fuentes I, Stegemann S, Golczyk H, Karcher D, Bock R (2014) Horizontal genome transfer as an asexual path to the formation of new species. Nature 511:232–235. https://doi.org/10.1038/nature13291Article Google Scholar
- Geraskin SA, Dikarev VG, Zyablitskaya YY, Oudalova AA, Spirin YV, Alexakhin RM (2003) Genetic consequences of radioactive contamination by the Chernobyl fallout to agricultural crops. J Environ Radioact 66:155–169. https://doi.org/10.1016/S0265-931X(02)00121-2Article Google Scholar
- Ghosh T, Anderson S, Elvidge C, Sutton P (2013) Using nighttime satellite imagery as a proxy measure of human well-being. Sustainability 5:4988–5019. https://doi.org/10.3390/su5124988Article Google Scholar
- Gillings MR, Stokes HW (2012) Are humans increasing bacterial evolvability? Trends Ecol Evol 27:346–352. https://doi.org/10.1016/j.tree.2012.02.006Article Google Scholar
- Grant PR, Grant BR (1992) Hybridization of bird species. Science 256:193–197. https://doi.org/10.1126/science.256.5054.193Article Google Scholar
- Hamilton C (2016) The Anthropocene as rupture. Anthr Rev 3:93–106. https://doi.org/10.1177/2053019616634741Article Google Scholar
- Hendry AP, Gotanda KM, Svensson EI (2017) Human influences on evolution, and the ecological and societal consequences. Philos Trans R Soc B 372:20160028. https://doi.org/10.1098/rstb.2016.0028Article Google Scholar
- Hodgson D, McDonald JL, Hosken DJ (2015) What do you mean, ‘resilient’? Trends Ecol Evol 30:503–506. https://doi.org/10.1016/j.tree.2015.06.010Article Google Scholar
- Huxel GR (1999) Rapid displacement of native species by invasive species: effects of hybridization. Biol Conserv 89:143–152. https://doi.org/10.1016/S0006-3207(98)00153-0Article Google Scholar
- Johnson MTJ, Munshi-South J (2017) Evolution of life in urban environments. Science 358:eaam8327. https://doi.org/10.1126/science.aam8327Article Google Scholar
- Kalusová V, Chytrý M, van Kleunen M, Mucina L, Dawson W et al (2017) Naturalization of European plants on other continents: the role of donor habitats. Proc Natl Acad Sci USA 114:13756–13761. https://doi.org/10.1073/pnas.1705487114Article Google Scholar
- Kost TD, Gessler C, Jänsch M, Flachowsky H, Patocchi A, Broggini GAL (2015) Development of the first cisgenic apple with increased resistance to fire blight. PLoS ONE 10:e0143980. https://doi.org/10.1371/journal.pone.0143980Article Google Scholar
- Laland KN, Uller T, Feldman MW, Sterelny K, Müller GB et al (2015) The extended evolutionary synthesis: its structure, assumptions and predictions. Proc R Soc B 282:20151019. https://doi.org/10.1098/rspb.2015.1019Article Google Scholar
- Lewis SL, Maslin MA (2015) Defining the Anthropocene. Nature 519:171–180. https://doi.org/10.1038/nature14258Article Google Scholar
- Liu J, Dietz T, Carpenter SR, Alberti M, Folke C et al (2007) Complexity of coupled human and natural systems. Science 317:1513–1516. https://doi.org/10.1126/science.1144004Article Google Scholar
- Lodge DM (1993) Biological invasions: lessons for ecology. Trends Ecol Evol 8:133–137. https://doi.org/10.1016/0169-5347(93)90025-kArticle Google Scholar
- Losey JE, Rayor LS, Carter ME (1999) Transgenic pollen harms monarch larvae. Nature 399:214. https://doi.org/10.1038/20338Article Google Scholar
- Ma JK-C, Barros E, Bock R, Christou P, Dale PJ et al (2005) Molecular farming for new drugs and vaccines. EMBO Rep 6:593–599. https://doi.org/10.1038/sj.embor.7400470Article Google Scholar
- Mallet J (2005) Hybridization as an invasion of the genome. Trends in Ecology Evolution 20:229–237. https://doi.org/10.1016/j.tree.2005.02.010Article Google Scholar
- Mallet J (2007) Hybrid Speciation. Nature 446:279–283. https://doi.org/10.1038/nature05706Article Google Scholar
- Malukiewicz J, Boere V, Fuzessy LF, Grativol AD, de Oliveira e Silva I et al (2015) Natural and anthropogenic hybridization in two species of eastern Brazilian marmosets (Callithrix jacchus and C. penicillata). PLoS ONE 10:e0127268. https://doi.org/10.1371/journal.pone.0127268Article Google Scholar
- McClure SB (2013) Domesticated animals and biodiversity: early agriculture at the gates of Europe and long-term ecological consequences. Anthropocene 4:57–68. https://doi.org/10.1016/j.ancene.2013.11.001Article Google Scholar
- McKinney ML (2006) Urbanization as a major cause of biotic homogenization. Biol Conserv 127:247–260. https://doi.org/10.1016/j.biocon.2005.09.005Article Google Scholar
- McKinney ML, Lockwood JL (1999) Biotic homogenization: a few winners replacing many losers in the next mass extinction. Trends Ecol Evol 14:450–453. https://doi.org/10.1016/S0169-5347(99)01679-1Article Google Scholar
- Milla R, Osborne CP, Turcotte MM, Violle C (2015) Plant domestication through an ecological lens. Trends Ecol Evol 30:463–469. https://doi.org/10.1016/j.tree.2015.06.006Article Google Scholar
- Morse NB, Pellissier PA, Cianciola EN, Brereton RL, Sullivan MM et al (2014) Novel ecosystems in the Anthropocene: a revision of the novel ecosystem concept for pragmatic applications. Ecol Soc 19:art12. https://doi.org/10.5751/es-06192-190212Article Google Scholar
- Muhlfeld CC, Kovach RP, Jones LA, Al-Chokhachy RK, Boyer MC et al (2014) Invasive hybridization in a threatened species is accelerated by climate change. Nat Clim Change 4:620–624. https://doi.org/10.1038/nclimate2252Article Google Scholar
- Nadeau CP, Urban MC, Bridle JR (2017) Climates past, present, and yet-to-come shape climate change vulnerabilities. Trends Ecol Evol 32:786–800. https://doi.org/10.1016/j.tree.2017.07.012Article Google Scholar
- Otto SP (2007) The evolutionary consequences of polyploidy. Cell 131:452–462. https://doi.org/10.1016/j.cell.2007.10.022Article Google Scholar
- Otto SP (2018) Adaptation, speciation and extinction in the Anthropocene. Proc R Soc B 285:20182047. https://doi.org/10.1098/rspb.2018.2047Article Google Scholar
- Pelletier F, Coltman DW (2018) Will human influences on evolutionary dynamics in the wild pervade the Anthropocene? BMC Biol 16:7. https://doi.org/10.1186/s12915-017-0476-1Article Google Scholar
- Pigeon G, Festa-Bianchet M, Coltman DW, Pelletier F (2016) Intense selective hunting leads to artificial evolution in horn size. Evol Appl 9:521–530. https://doi.org/10.1111/eva.12358Article Google Scholar
- Pimentel D, McNair S, Janecka J, Wightman J, Simmonds C et al (2001) Economic and environmental threats of alien plant, animal, and microbe invasions. Agricult Ecosyst Environ 84:1–20. https://doi.org/10.1016/S0167-8809(00)00178-XArticle Google Scholar
- Potts SG, Biesmeijer JC, Kremen C, Neumann P, Schweiger O, Kunin WE (2010) Global pollinator declines: trends, impacts and drivers. Trends Ecol Evol 25:345–353. https://doi.org/10.1016/j.tree.2010.01.007Article Google Scholar
- Rhymer JM, Simberloff D (1996) Extinction by hybridization and introgression. Annu Rev Ecol Syst 27:83–109. https://doi.org/10.1146/annurev.ecolsys.27.1.83Article Google Scholar
- Richardson DM, Pyšek P, Rejmánek M, Barbour MG, Panetta FD, West CJ (2000) Naturalization and invasion of alien plants: concepts and definitions. Divers Distrib 6:93–107. https://doi.org/10.1046/j.1472-4642.2000.00083.xArticle Google Scholar
- Rudman SM, Kreitzman M, Chan KMA, Schluter D (2017) Evosystem services: rapid evolution and the provision of ecosystem services. Trends Ecol Evol 32:403–415. https://doi.org/10.1016/j.tree.2017.02.019Article Google Scholar
- Schumer M, Cui R, Rosenthal GG, Andolfatto P (2015) Reproductive isolation of hybrid populations driven by genetic incompatibilities. PLoS Genet 11:e1005041. https://doi.org/10.1371/journal.pgen.1005041Article Google Scholar
- Simberloff D, Martin J-L, Genovesi P, Maris V, Wardle DA et al (2013) Impacts of biological invasions: what’s what and the way forward. Trends Ecol Evol 28:58–66. https://doi.org/10.1016/j.tree.2012.07.013Article Google Scholar
- Simmonds JS, Watson JEM, Salazar A, Maron M (2019) A composite measure of habitat loss for entire assemblages of species. Conserv Biol 5:4. https://doi.org/10.1111/cobi.13331Article Google Scholar
- Song Q, Zhang T, Stelly DM, Chen ZJ (2017) Epigenomic and functional analyses reveal roles of epialleles in the loss of photoperiod sensitivity during domestication of allotetraploid cottons. Genome Biol 18:99. https://doi.org/10.1186/s13059-017-1229-8Article Google Scholar
- Soucy SM, Huang J, Gogarten JP (2015) Horizontal gene transfer: building the web of life. Nat Rev Genet 16:472–482Article Google Scholar
- Steffen W, Crutzen J, McNeill JR (2007) The Anthropocene: are humans now overwhelming the great forces of Nature? Ambio 36:614–621Article Google Scholar
- Steffen W, Persson Å, Deutsch L, Zalasiewicz J, Williams M et al (2011) The Anthropocene: from global change to planetary stewardship. Ambio 40:739–761. https://doi.org/10.1007/s13280-011-0185-xArticle Google Scholar
- Steffen W, Richardson K, Rockström J, Cornell SE, Fetzer I et al (2015) Planetary boundaries: guiding human development on a changing planet. Science 347:1259855. https://doi.org/10.1126/science.1259855Article Google Scholar
- Stephens AEA, Krannitz PG, Myers JH (2009) Plant community changes after the reduction of an invasive rangeland weed, diffuse knapweed, Centaurea diffusa. Biol Control 51:140–146. https://doi.org/10.1016/j.biocontrol.2009.06.015Article Google Scholar
- Sullivan AP, Bird DW, Perry GH (2017) Human behaviour as a long-term ecological driver of non-human evolution. Nat Ecol Evol 1:1–11. https://doi.org/10.1038/s41559-016-0065Article Google Scholar
- Thomas CD (2013) The Anthropocene could raise biological diversity. Nature 502:7. https://doi.org/10.1038/502007aArticle Google Scholar
- Thompson JN (1999) The evolution of species interactions. Science 284:2116–2118. https://doi.org/10.1126/science.284.5423.2116Article Google Scholar
- Toussaint A, Beauchard O, Oberdorff T, Brosse S, Villéger S (2016) Worldwide freshwater fish homogenization is driven by a few widespread non-native species. Biol Invas 18:1295–1304. https://doi.org/10.1007/s10530-016-1067-8Article Google Scholar
- Van Eenennaam AL (2017) Genetic modification of food animals. Curr Opin Biotechnol 44:27–34Article Google Scholar
- van Nes EH, Arani BMS, Staal A, van der Bolt B, Flores BM et al (2016) What do you mean, ‘tipping point’? Trends Ecol Evol 31:902–904. https://doi.org/10.1016/j.tree.2016.09.011Article Google Scholar
- Vanwalleghem T, Gómez JA, Infante Amate J, González de Molina M, Vanderlinden K et al (2017) Impact of historical land use and soil management change on soil erosion and agricultural sustainability during the Anthropocene. Anthropocene 17:13–29. https://doi.org/10.1016/j.ancene.2017.01.002Article Google Scholar
- Vitousek PM (1997) Human domination of earth’s ecosystems. Science 277:494–499. https://doi.org/10.1126/science.277.5325.494Article Google Scholar
- Vitousek PM, Naylor R, Crews T, David MB, Drinkwater LE et al (2009) Nutrient imbalances in agricultural development. Science 324:1519–1520. https://doi.org/10.1126/science.1170261Article Google Scholar
- Walther G-R, Roques A, Hulme PE, Sykes MT, Pysek P et al (2009) Alien species in a warmer world: risks and opportunities. Trends Ecol Evol 24:686–693. https://doi.org/10.1016/j.tree.2009.06.008Article Google Scholar
- Wolfenbarger LL, Phifer PR (2000) The ecological risks and benefits of genetically engineered plants. Science 290:2088–2093. https://doi.org/10.1126/science.290.5499.2088Article Google Scholar
- Worm B, Paine RT (2016) Humans as a hyperkeystone species. Trends Ecol Evol 31:600–607. https://doi.org/10.1016/j.tree.2016.05.008Article Google Scholar
- Zalasiewicz J, Williams M, Waters CN, Barnosky AD, Palmesino J et al (2017) Scale and diversity of the physical technosphere: a geological perspective. Anthropocene Rev 4:9–22. https://doi.org/10.1177/2053019616677743Article Google Scholar
- Zhang C, Wohlhueter R, Zhang H (2016) Genetically modified foods: a critical review of their promise and problems. Food Sci Hum Wellness 5:116–123. https://doi.org/10.1016/j.fshw.2016.04.002Article Google Scholar
Acknowledgements
Financial support was provided by the Ministério do Meio Ambiente/PROBIO II. We thank Professor Stuart A. Newman, Deborah Klosky, and one anonymous reviewer for comments on the manuscript, and Mr. Roy Funch for linguistic advice.
Author information
Affiliations
- Instituto de Pesquisas Jardim Botânico do Rio de Janeiro, Rio de Janeiro, BrazilPablo José Francisco Pena Rodrigues & Catarina Fonseca Lira
Corresponding author
Correspondence to Pablo José Francisco Pena Rodrigues.
